素粒子から宇宙の起源に至る広範な事象の説明を可能にした物理学の成果には、ただただ驚嘆するばかりです。生物も物質から出来ている以上、物理法則ですべてを説明できるはずだという考えも分かりますが、それでは生物学を生業(なりわい)とする研究者としての基盤そのものが怪しくなる気がします。数式が不得意だと言う理由で生物学科に進んだ私としましては、生物を支配する未知の法則が眠っていて、あえて難しい物理の理論や数学を使わなくてもそれらを発見できると信じてきました。勿論、その法則は生気論(生き物には魂や“気”が宿るという考え)の類ではなくて、メンデルの遺伝法則のように、最終的には物理学の言葉で置き換えられるべきものであり、当然、近代生物学の知識と矛盾するものであってはなりません。
【自己複製と物理学】
ここでは、生物の最大の特性であります≪自己複製≫(分裂)に注目します。今回のコラムを書くに当たり、友人である複数の物性物理学者と話して確認したのですが、物理学では自己複製する系を取り扱った経験がない、ということはどうも確かなようです(むしろ、「目に見える物体から素粒子に至るまで、生物を除いて自己複製するものはない」と言うべきでしょう)。つまり、自己複製系は基礎物理学者にとって“聖域”ということになりますから、宝物が隠されているかも知れません。さて、その宝物を見つけ出す手段ですが、例によって瞑想だけが頼りです。「自然は単純で美しい」と言う、物理学者の言葉を一途に信じて、できるだけ単純で美しいモデルを作ることにしました。
【キーワードは元本保証】
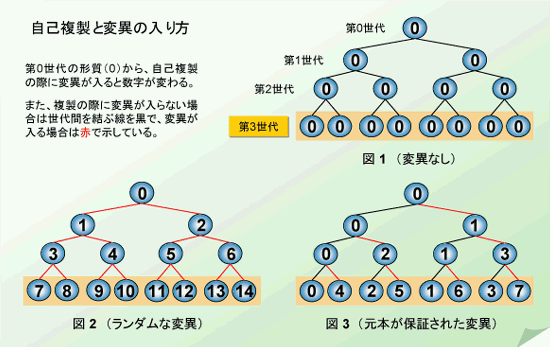
まず、下に示した図のうちの図1をご覧ください。これは概念図ですから、○は自己複製をするユニットであれば何でもよく、DNA・遺伝子・ウイルス・バクテリア(細菌)・真菌類(かび・酵母など)・培養細胞・個体、場合によっては、種(しゅ)でも、あるいは文化のようなものでもかまいません。分かり易くするために、図には第3世代までしか家系図が描かれていません。四角で囲まれた最下段の8個だけがが現存する実体(第3世代)です。つまり、それぞれのユニットが自己複製を終えると2個の子供が出来ますが、その親は自動的に消滅します。図1は細菌の複製様式そのものですから、これ以上何も付け加える必要がなさそうですが、実はこの集団の存続はあやふやなのです。何故なら、どの細菌も同じ表現型(形や機能)(0)ですから、もし偶然抗生物質に接したとしますと、耐性がない場合全滅してしまいます。この悲劇から逃れる術は一つしかありません。それは、前もって表現型の多様性を確保しておくことです。うまくすれば、その抗生物質に耐性の個体が準備されているかも知れません。これがダーウィン進化の基本的な考え方です。
そこで、表現型の変化を考慮したのが図2です。ここでは自己複製の度に、親と違った表現型をもつ2個体の子供が出来るようにしてあります。(数字の違いは表現型が異なることを意味します。図間の数字は関係がありません。)現在生きている細菌は最下段の7、8、9、10、11、12、13、14と番号を振った8個体です。多士済済のメンバー構成ですが、一見して、この集団も極めて危険な状況にあることが分かります。と言うのは、もし、現在の環境が永い間変わらなかったとしますと、先祖の0型が居ませんので、全滅する危惧があります。一方、幸運にも10番の細菌が抗生物質に耐性を持っており、生き延びたとしても、次の世代では必ず新しい表現型が加わりますので、将来、耐性が保持されるかどうかは「神のみぞ知る」です。では、この問題をどのように解決すればいいのでしょう。いろいろな答えがあるでしょうが、私が一番“単純で美しい”と思う解を示しましょう。
図3を見てください。ここでは、それぞれのユニットが自己複製して生ずる2個体の子供のうち、一方は必ず親と同じ表現型を持つようにしてあります。現存する8匹の細菌の表現型は0、1、2、3、4、5、6、7まで、見事に全部そろっています。このモデルの特徴は、1)先祖型の表現型(0)は何世代経っても保証される、2)過去に一度現れた表現型は、その個体が偶然死滅しない限りいつまでも存在し続ける、3)表現型が一度に極端に変わっても、原理的には全滅しない、の3つです。一口で表現しますと、「元本保証の多様性創出モデル」です。一種の“不等分裂”とも言うべきこの方式ですと、次々と違った環境(選択圧)に曝されても、適応進化して生き延びて行けそうです。
|